Xavier Morin
Division cellulaire et neurogenèse
Contexte
Toutes les cellules nerveuses, neurones et cellules gliales, produites au cours du développement du système nerveux des Vertébrés dérivent d’un réservoir initial de cellules souches organisées en une monocouche cellulaire, le neuroepithelium. Après une étape initiale de prolifération de ce réservoir, grâce à des divisions symétriques (mitoses), apparaissent des divisions dites asymétriques qui produisent deux cellules filles différentes : l’une demeure une cellule souche (auto renouvellement) tandis que l’autre va se différencier en neurone. La régulation spatio-temporelle de la transition divisions symétriques prolifératives versus asymétriques neurogéniques détermine le moment d’apparition, la position des différentes populations de neurones et in fine la taille du cerveau. Les mécanismes qui président au choix entre ces deux modes de division ne sont pas encore bien compris. L’enjeu est important au niveau fondamental, mais aussi pour mieux comprendre l’origine de certaines pathologies, comme des microcéphalies.
Cette problématique est au cœur des recherches développées par l’équipe qui étudie la division asymétrique, processus générateur de diversité cellulaire, chez les vertébrés et les invertébrés.
Résultats marquants
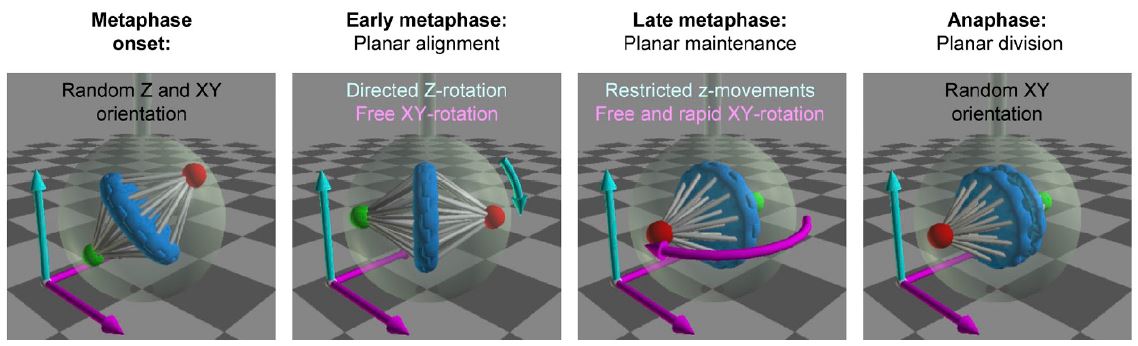
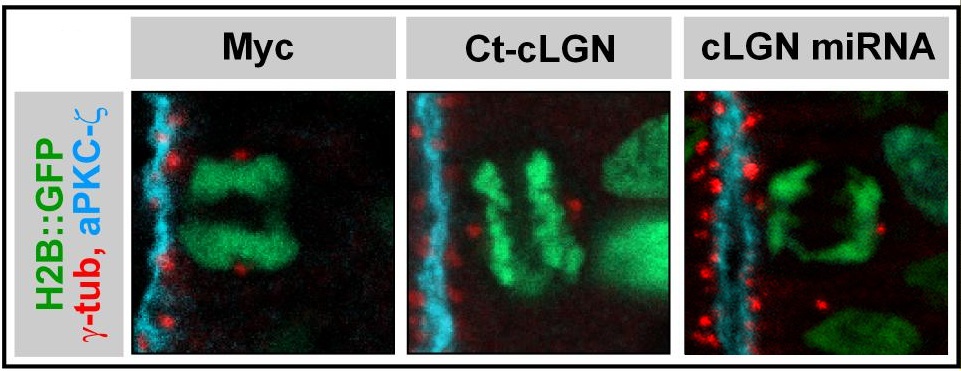
Dans de nombreux modèles cellulaires, les choix d’identité « asymétriques » entre cellules sœurs sont corrélés à l’orientation de l’axe de division de la cellule mère dans le tissu, ce qui a mené à l’idée que l’orientation « contrôle » le destin des cellules filles. Cette hypothèse nous a incités à explorer les mécanismes qui contrôlent l’orientation des divisions cellulaires, et nous avons ainsi contribué à identifier et caractériser une cascade moléculaire largement conservée au cours de l’évolution et mise à profit aussi bien dans des divisions symétriques qu’asymétriques (Morin et al, 2007 ; Peyre et al, 2011, Saadaoui et al, 2014, 2017 ; di Pietro et al, 2017). En revanche, l’étude des défauts développementaux associés à une perturbation de l’axe de division des progéniteurs neuraux montre qu’elle n’a pas ou peu d’effet direct sur l’acquisition de l’identité des cellules filles (cellule souche ou neurone). Il semblerait que, tout au moins dans le neuroépithélium de la moelle épinière, sur laquelle nous menons l’essentiel de nos travaux, l’orientation planaire du fuseau mitotique réponde principalement à la nécessité de préserver l’architecture de ce tissu en y maintenant les cellules filles issues des mitoses (Morin et al, 2007).
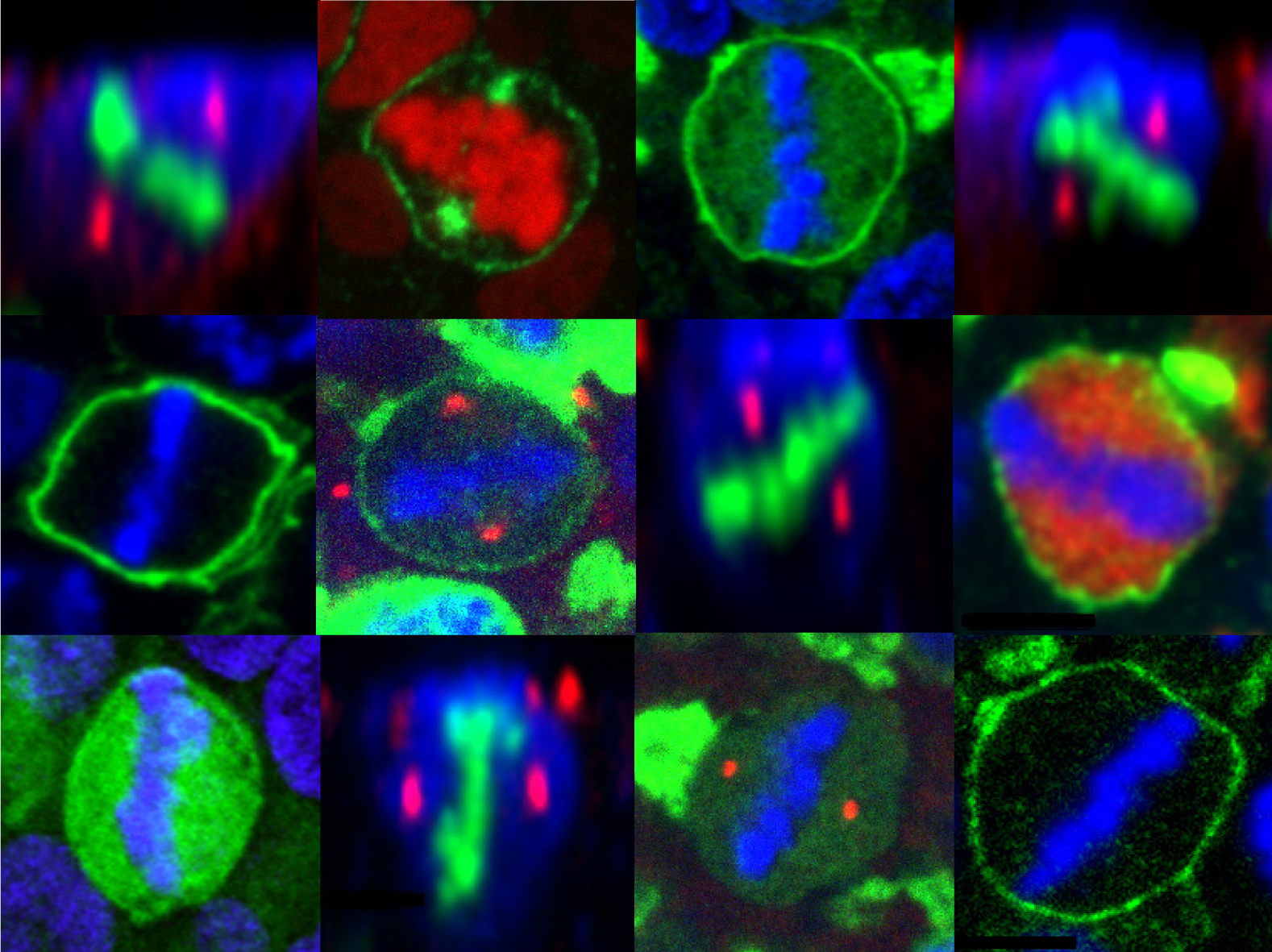
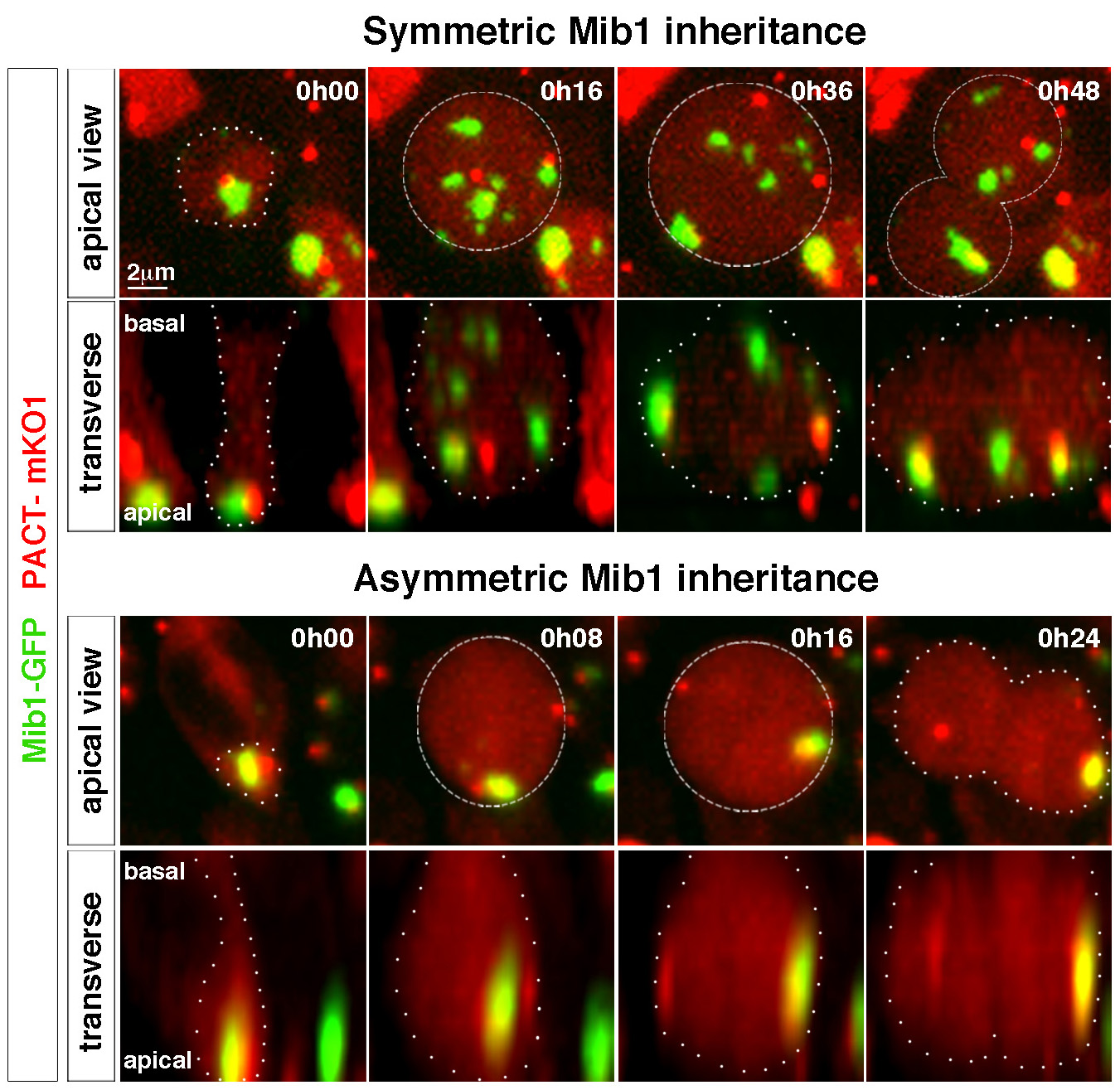
En parallèle, nous avons donc exploré d’autres mécanismes qui pourraient contrôler l’asymétrie de division dans les cellules souches neurales. Ainsi, nous avons récemment observé la distribution asymétrique d’un régulateur de la voie de signalisation Notch, l’ubiquitine ligase Mindbomb (Mib1) dans les progéniteurs neuroepithéliaux en division. Mib1, associé à une organelle encore mal connue, le satellite pericentriolaire, est enrichi auprès du centriole le plus « jeune » en interphase, ce qui aboutit à sa localisation à un seul pôle du fuseau mitotique et à son héritage par une seule cellule fille qui deviendra alors un neurone après la division cellulaire. En combinant des gains et pertes de fonction et l’imagerie en temps réel de la division et de la différentiation cellulaires dans le tube neural de poulet, nous avons montré que cette asymétrie de distribution de Mib1 est instructive pour l’identité des cellules sœurs. Plus remarquable encore, nous avons aussi montré qu’une partie des progéniteurs « échappe » à cette asymétrie grâce à la présence d’une réserve supplémentaire de Mib1 stockée au niveau de l’appareil de Golgi en interphase, et envoyée dans la cellule fille qui ne reçoit pas le pool centrosomal de Mib1 (Tozer et al, 2017). En accord avec une transition progressive entre les différents modes de division, nous avons observé que la localisation de Mib1 évolue entre la phase précoce de prolifération et les stades de neurogenèse plus tardifs.
Projets en cours
Ces résultats sont particulièrement intrigants et nous mènent vers d’autres questions. Comment Mib1 est-il recruté au Golgi, et quels sont les partenaires moléculaires qui régulent le trafic de Mib1 entre Golgi et satellites centriolaires pour contrôler les choix d’identité ? Un rôle connu de Mib1 est de modifier Dll1, un ligand de Notch, à la membrane cellulaire, et c’est vraisemblablement là que Mib1 est actif, le Golgi et les satellites centriolaires remplissant essentiellement un rôle de stockage et de routage. Quand et comment Mib1 transite-t-il entre satellites et membrane plasmique ? Répondre à ces questions nécessite un suivi à haute résolution de la localisation sub-cellulaire de Mib1 au cours du temps, à la fois à l’échelle du cycle cellulaire, mais aussi au cours du temps développemental. Il est également nécessaire de pouvoir isoler Mib1 pour identifier ses partenaires spécifiques dans différents compartiments cellulaires. Nous développons actuellement les outils nécessaires pour y répondre.
A un même stade développemental dans le neuroepithelium, on observe simultanément des divisions symétriques et asymétriques, mais au sein d’un même clone cellulaire, les deux modes de division se succèdent et la transition entre divisions symétriques et asymétrique semble irréversible, suggérant que le choix du mode de division n’est pas stochastique mais déterminé. Au-delà de la question du contrôle de la mise en place de l’asymétrie qui permettra l’acquisition d’une identité différente entre cellules filles suite à une division neurogenique, ceci implique qu’un choix en amont, sous contrôle de programmes génétiques spécifiques, détermine le mode de division des progéniteurs entre prolifératif (symétrique) et neurogénique (asymétrique). Cette question, qui a été peu abordée dans les modèles « historiques » de division asymétrique basés sur des lignages invariants chez les invertébrés, est au cœur d’un nouveau projet développé dans l’équipe, qui vise à identifier ces programmes en étudiant les différences de profil transcriptionnel et d’organisation de la chromatine entre progéniteurs en mode de division symétrique versus asymétrique.
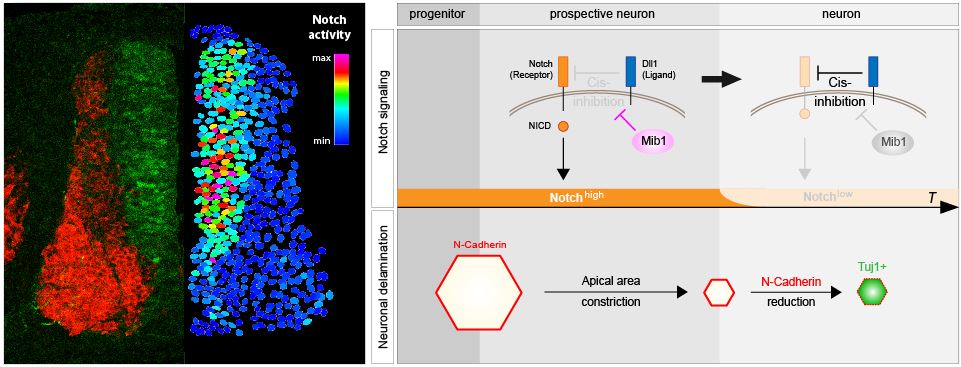
Baek C, Freem L, Goïame R, Sang H, Morin X, Tozer S. Mib1 prevents Notch Cis-inhibition to defer differentiation and preserve neuroepithelial integrity during neural delamination.
PLoS Biology 2018, 16(4):e2004162.
di Pietro F, Valon L, Li Y, Goïame R, Genovesio A, Morin X. An RNAi screen in a novel model of oriented divisions identifies the actin capping protein Z β as an essential regulator of spindle orientation.
Current Biology 2017, 27(16):2452–2464
Saadaoui M, Konno D, Loulier K, Goiame R, Jadhav V, Mapelli M, Matsuzaki F, Morin X. Loss of the canonical spindle orientation function in the Pins/LGN ortholog AGS3.
EMBO Reports 2017, 18(9):1509-1520
Tozer S, Baek C, Fischer E, Goïame R, Morin X. Differential routing of Mindbomb1 via centriolar satellites regulates asymmetric divisions of neural progenitors.
Neuron 2017, 93(3):542-551.
Saadaoui M, Machicoane M, di Pietro F, Etoc F, Echard A, Morin X. Dlg1 controls planar spindle orientation in the neuroepithelium through direct interaction with LGN.
The Journal of Cell Biology 2014, 206(6):707-717.
Loulier K, Barry R, Mahou P, Le Franc Y, Supatto W, Matho KS, Siohoi I, Fouquet S, Dupin E, Benosman R, Chedotal A, Beaurepaire E, Morin X, Livet J. Multiplex cell and lineage tracking with combinatorial labels.
Neuron 2014, 81(3):505-520.